Primary Mechanistic Interpretation
The most compelling Sphenocentrum hypothesis after de-risking is combined steroid-metabolism and endocrine- enzyme modulation rather than direct androgen receptor agonism or PDE5A-driven erectile pharmacology.
Aromatase/CYP19A1 is the cleanest structural efficacy hypothesis for pinnatasterone. It is reference-pocket aligned
(~2.8 Å from the testosterone co-crystal subpocket) with favorable anchor contacts. 20-hydroxyecdysone and isocolumbin also align on CYP19A1. This fits a testosterone-to-estrogen conversion axis upstream of receptor activation and parallels the aromatase emphasis in the Tongkat literature [1,2].
The figure below shows the quantum-computed electrostatic basis for this selectivity. Pinnatasterone carries electron-rich oxygen groups with computed partial charges of approximately −0.38 to −0.45 e. These groups are geometrically and electrostatically complementary to ARG115, the positively charged guanidinium residue in the CYP19A1 binding pocket, forming the same four anchor contacts (ARG115, PHE134, PHE221, TRP224) as the approved aromatase inhibitor exemestane. The androgen receptor, by contrast, presents an entirely hydrophobic binding core (PHE764, PHE876, PHE891, TRP741) with no ionic partner for the electron-rich oxygens, explaining the weaker predicted binding (2/4 contacts, −6.70 kcal/mol vs −8.97 kcal/mol on CYP19A1).
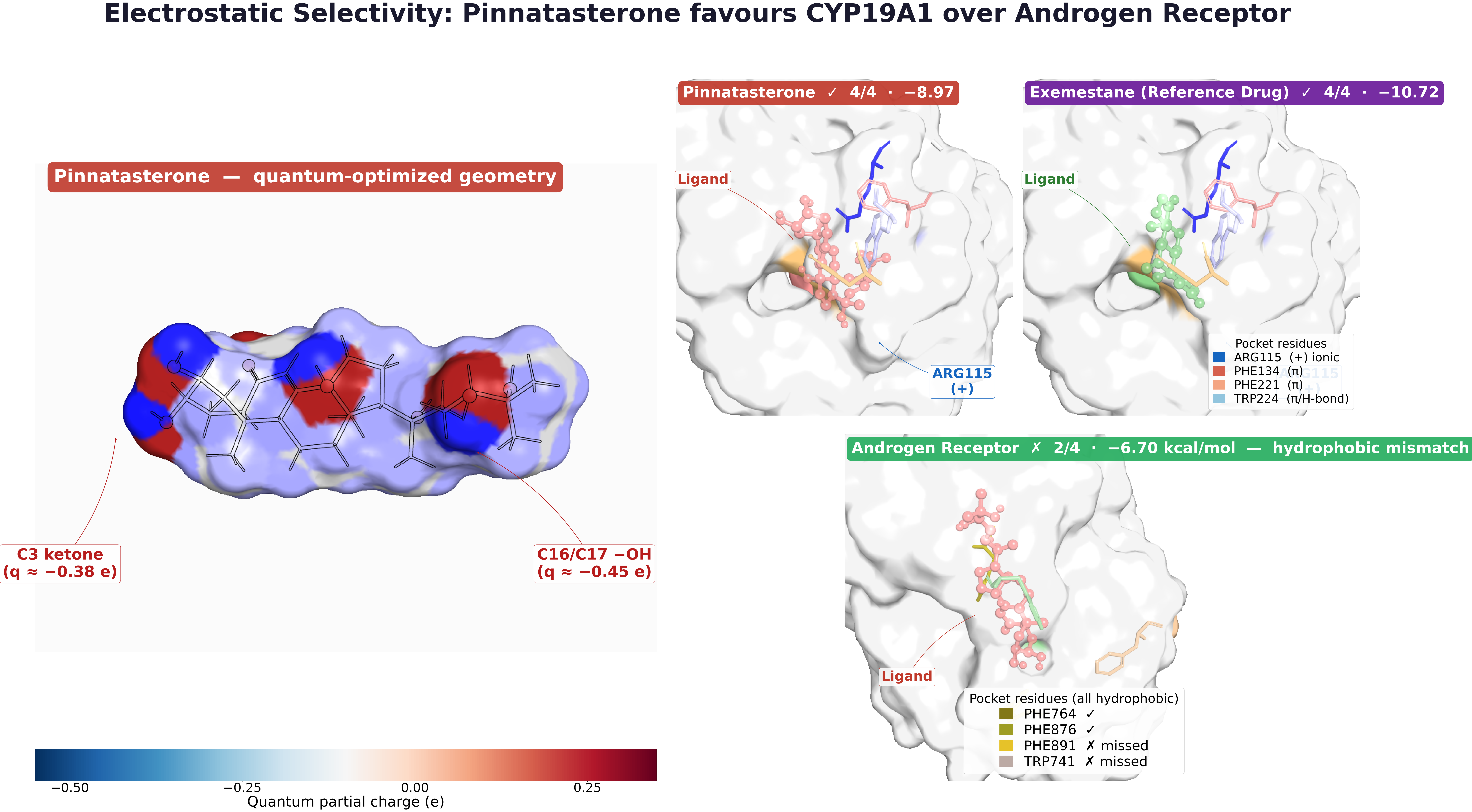
Electrostatic selectivity panel: pinnatasterone quantum-optimized geometry with partial-charge surface (left), compared to its binding pose in CYP19A1 alongside exemestane reference drug (top right) and the androgen receptor (bottom right).
SRD5A2 provides the second mechanistic route. SRD5A2 controls conversion of testosterone to dihydrotestosterone. Isocolumbin achieved the strongest docking score on SRD5A2, but static de-risking classifies it as partial (~6.6 Å from the finasteride reference subpocket). Columbamine is the best reference-aligned
Sphenocentrum marker on SRD5A2 (~4.9 Å). Assay planning should prioritize columbamine for structural plausibility and include isocolumbin as a docking-led follow-up with explicit pose uncertainty.
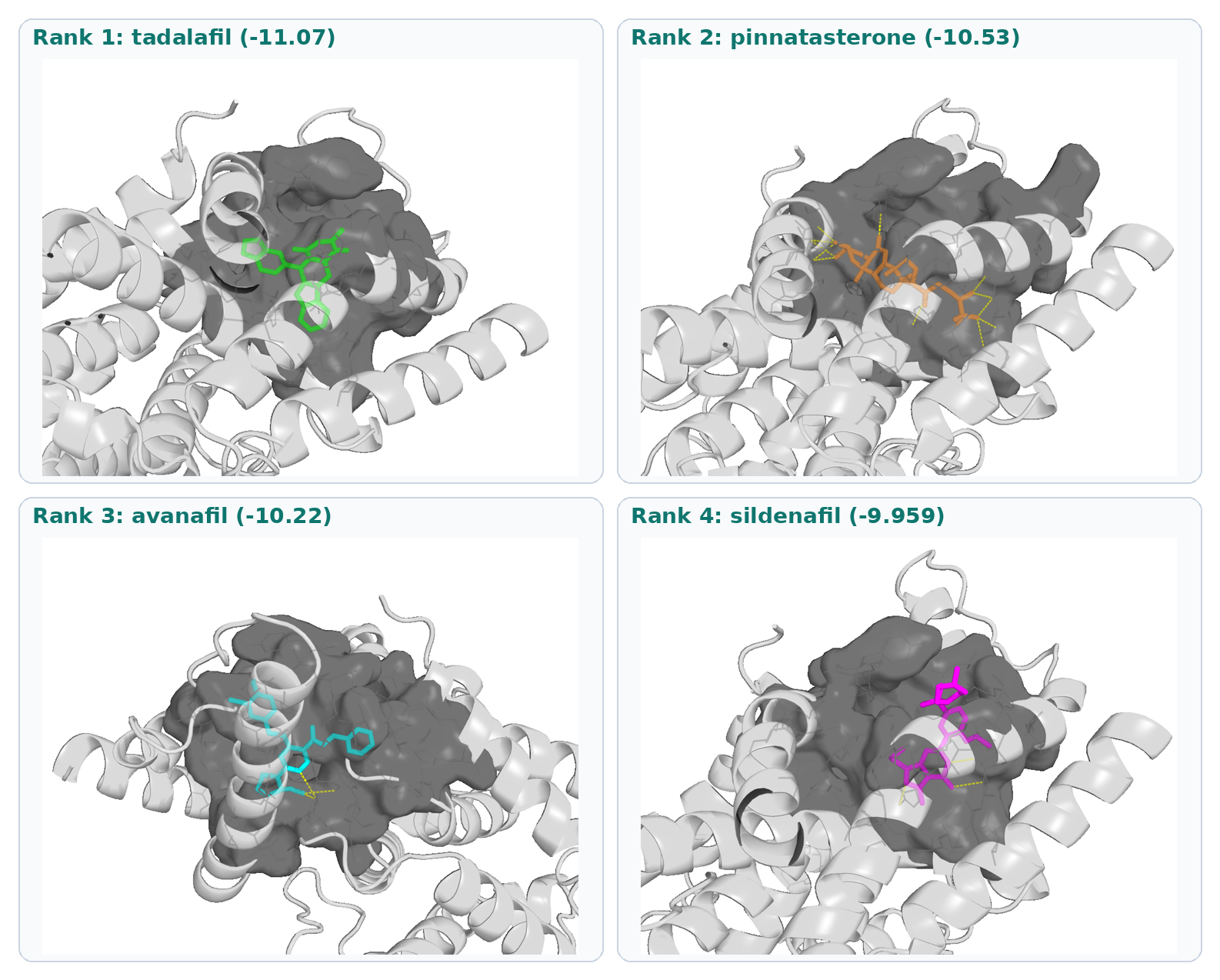
The PDE5A signal is a cautionary example of score-versus-mode divergence. Pinnatasterone scored below tadalafil
but above several approved PDE5 drugs in molecular docking, yet it does not reproduce the inhibitor pharmacophore/subpocket geometry that tadalafil and the co-crystal ligand satisfy. That pattern is common when large phytoactive scaffolds dock in the broader catalytic pocket without matching the drug binding mode. Erectilefunction claims should not rest on this target unless biochemistry contradicts the structural result.
CYP17A1 showed favorable docking scores for pinnatasterone but reference-centroid distances near ~29 Å, indicating off-pocket or unreliable poses under the current binding-site setup. Androgen-biosynthesis interpretation from CYP17A1 should wait on pose repair or orthogonal modeling.
The androgen receptor remains a secondary context. Columbin and several alkaloids show reference-aligned or near-aligned poses in the testosterone pocket, but steroid controls still dominate molecular docking. Direct AR agonism is not the primary story.
The PXR signal is best interpreted as a safety finding. PXR activation can induce drug-metabolizing enzymes and transporters. Strong docking by isocolumbin and related markers, together with mixed static alignment on PXR, indicates that a standardized, high-bioavailability extract concept would need CYP induction and drug-interaction evaluation before any product-facing claims.